Ochnaceae - Ochnaceae
| Ochnaceae | |
|---|---|
 | |
| Sauvagesia erecta из южной Бразилии | |
| Научная классификация | |
| Королевство: | Plantae |
| Clade: | Трахеофиты |
| Clade: | Покрытосеменные |
| Clade: | Eudicots |
| Clade: | Росиды |
| Заказ: | Мальпигиалес |
| Семья: | Ochnaceae ОКРУГ КОЛУМБИЯ.[1] |
| Роды | |
Видеть текст | |
Ochnaceae это семья из цветущие растения в порядок Мальпигиалес.[2] в Система APG III из классификация из цветковых растений, Ochnaceae определяется в широком смысле и включает около 550 разновидность,[3] и охватывает то, что некоторые систематики рассматривали как отдельный семьи Medusagynaceae и Quiinaceae.[1] В филогенетический изучать что было опубликовано в 2014 году Ochnaceae получил широкое признание смысл,[4] но два работает опубликовано после того, как APG III приняли небольшие семейства Medusagynaceae и Quiinaceae.[3][5] Они не были приняты APG IV (2016).
В этой статье «Ochnaceae» будет относиться к более крупным ограничение из семейства, иначе известного как Ochnaceae Sensu lato или как очноиды.[6] В этом смысле семейство включает 32 рода, насчитывающих около 550 видов.[7]
Ochnaceae, в широком или узком определении, - это пантропический в распределение, с несколькими видами культивируется вне этого классифицировать. Ochnaceae наиболее разнообразный в неотропы, со вторым центр разнообразия в тропическая африка.[4] Он состоит в основном из кустарников и небольших деревьев, а в Sauvagesia, немного травянистый разновидность. Многие из них - деревца, с одиночными прямостоячими хобот, но невысокого роста. Ochnaceae отличаются необычными листьями. Обычно они блестящие, с близко расположенными параллельными жилками, зубчатыми краями и заметными. прилистники. Большинство видов жужжание опылено.[8] У восьми родов в племя Sauvagesieae, цветок меняется форма после открытие, продолжение рост из ткань внутри цветка.[4]
Несколько видов Очна находятся культивируется в качестве украшения.[9] Очна томасиана вероятно, наиболее часто посажен, но это часто неправильно идентифицированный в садоводстве литература.[10]
Листья Цеспедезия иногда до 1 м (3,3 фута) в длина и используются для кровля.[11] An травяной чай сделано из пантропика сорняк Sauvagesia erecta.
В своем эволюция, Ochnaceae был необычным, в "возвращение " к состояния персонажей которые рассматриваются как наследственный или же примитивный. Например, актиноморфный цветочная симметрия дважды появлялся в подсемейство Ochnoideae. Также два клады из Ochnaceae, один у Ochnoideae и другой у Quiinoideae полученный состояние очень близко к апокарпия. Полное разделение плодолистики (апокарпия ) считается родовое государство за покрытосеменные.[12]
Окаменелости относимые к Ochnaceae, известны с ранних эоцен из Миссисипи.[13] В возраст семьи очень грубо по оценкам через 100 миллионов лет.[14]
Великое множество названия родов были опубликованы в Ochnaceae.[15] В таксономический при ревизии Ochnaceae, как трех семейств, в 2014 г. были приняты только 32 из этих родов; один у Medusagynaceae, четыре у Quiinaceae и 27 у Ochnaceae SS..[3] В том же году 33-й род, Некия, был восстановлен с целью сохранения монофилия другого рода, Sauvagesia.[4]
Самые крупные роды Ochnaceae: Урата (200 видов), Очна (85), Кампилоспермум (65), Sauvagesia (39), и Quiina (34).[3] Ни один из более крупных родов не был предметом филогенетический анализ из Последовательности ДНК избранных гены. В одном исследовании подсемейства Quiinoideae, основанном на trn L-F межгенный спейсер, только девять видов были отобранный из этого подсемейства.[16]
Роды
Следующий список из 33 родов состоит из Некия, который был воскрешен в 2014 году,[4] плюс 32 рода, которые были описанный в последней редакции Ochnaceae.[3][17][18] Классификация от Schneider. и другие (2014).[4]
|
|
Описание
Следующее описание является выдержка из описаний Medusagynaceae, Quiinaceae и Ochnaceae SS. в Семейства и роды сосудистых растений,[3][17][18] с некоторой информацией из других источников, как указано.
По большей части кусты и маленький деревья, плюс несколько деревьев среднего размера, а в Sauvagesia, немного травы.
Листья просто, Кроме перистый сложный в Krukoviella,[2] и у Quiinoideae, часто перисто дольчатый или соединение на малолетний растения. Перистые листья типичны для Ритидантера.[4] Листья часто бывают кожный и заметно зубчатый. Прилистники присутствует, кроме Медузагайн.
Жилкование довольно часто скалярный (лестничный) по внешнему виду, с параллельными и близко расположенными вторичный и высшее вены. Черешки отсутствующий или короткие, иногда напоминающие Pulvinus.[2]
Унисекс цветы распространены в Medusagyne и Quiinoideae (кроме Froesia), но ограничивается кладой из трех родов у Ochnoideae. Однополые цветы встречаются в Schuurmansia, Schuurmansiella, и Евфемида.[4] Цветы всегда однополые в Schuurmansiella.[3]
В полигамный виды, цветы оценены как бисексуал на основе морфология Только.[16] Пыльца произведено очевидно гермафродитный цветы в некоторых случаях оказывались невосприимчивый, рендеринг цветка функционально женский.
Чашелистики От 3 до 5, часто неравные, иногда нарастающий.
Лепестки 4 или 5 или редко 3, 6, 7 или 8, часто искажать, свободный или же плавленый только у основания, иногда загибается над чашелистиками.
Плодородный тычинки От 5 до 10 или многочисленные, редко один. Нити иногда настойчивый, иногда сужается возле пыльники.
Пыльники базисный или немного с тыльной стороны, обычно расчесывание на один или два апикальный или субапикальные поры, иногда неправильно к продольный прорези. В Медузагайн и Quiinoideae, увеличенный перегородка отделяет теки.
Стаминодес часто присутствует, бесплатно или родиться, иногда лепестковидная, иногда охватывающая плодородные тычинки.
Нектар нет произведено. Цветы обычно опыляемый жужжанием.
Яичник начальство, продольно ребристый в Медузагайн и Quiinoideae; без ребер у Ochnoideae. Carpels полностью слились или почти разделились; 2-15 или до 25 дюймов Медузагайн. Стиль апикальный или гинекологический.
Фрукты иногда крылатый; редко орех или же костянка, довольно часто ягода -подобно; обычно септицидный капсула, иначе яичник отделяется, образуя черноватый костянки на обычно красноватом, нарастающий сосуд.
Семена белковый или же великолепный крылатые или нет. В семенная оболочка часто включает слой кристаллические ячейки. Это склереиды, каждый из которых содержит оксалат кальция кристаллы в виде друза.
Классификация
Примерно до конца 20-го века Ochnaceae считались довольно странным семейством, которое трудно определить с высокой степенью уверенности. Даже в 21 веке некоторые авторы относился к роду Страсбургерия как ближайший родственник Ochnaceae, а некоторые даже поместили его в семью.[19] в Система APG III, Страсбургерия сгруппирован с Иксерба сформировать семью Strasburgeriaceae в розид порядок Crossosomatales.[20]
Реже род Дигодендрон считалось близким к Страсбургерия и Ochnaceae. Молекулярно-филогенетический исследования сильно поддерживается включение Дигодендрон в порядке розида Мальвалес, и это иногда рассматривается как моноспецифический семья в нем.[21] В противном случае он был помещен в Bixaceae, хотя есть основания подозревать, что он может быть ближе к Sphaerosepalaceae.[20]
Все упомянутые выше роды, а также Ochnaceae долгое время считались анамальный таксоны неопределенного близость. Все они в то или иное время помещались вместе с Ochnaceae, недалеко от Theaceae, семья теперь включена в базальный астерида порядок Ericales.[20]
В 2012 году анализ хлоропласт ДНК решено Ochnaceae как сестра к группа пяти семей, известных как кластиоиды.[5] Этот результат имел только слабые бутстрап поддерживать. Кластиоиды когда-то считались группой из четырех семейств,[22] но Clusiaceae был разделен в 2009 году[6] и имя Calophyllaceae был воскрешен для одного из образовавшихся сегрегаций.[1][23]
Есть всего несколько морфологический символы который объединяться кластероиды с Ochnaceae. Лепесток праздник часто искажать у кластиоидов и обычно так у Ochnaceae. В обеих группах цветы обычно несут многочисленные тычинки, а в яичник, то плацентация в основном подмышечный. в семяпочки, то нуцеллус часто бывает тонким, а внешний покров обычно толще внутреннего.[24]
Таксономия
Ochnaceae делится на три подсемейства: Medusagynoideae, Quiinoideae и Ochnoideae.[4]
Молекулярное филогенетическое исследование разрешило Medusagynoideae и Quiinoideae как сестра подсемейств, но этот результат имел только слабые статистическая поддержка.[5] Цветки обоих подсемейств полисистемный. Кроме рода Froesia, многие или все цветы однополый. В пыльники содержать массивный перегородка между теки который сохраняется после пыльника расхождение. В стили расходятся наружу от яичника. В цветение, яичник оформлен продольными ребрами.[25]
Medusagynoideae состоит из одного вида: Медузагина оппозитнолистная. это эндемичный к остров из Маэ в Сейшельские острова.[26]
Quiinoideae насчитывает около 48 видов четырех родов: Froesia, Quiina, Touroulia, и Лакунария. Это ограничено тропический Америка.[27] Froesia отличается от трех других родов. Его цветки всегда обоеполые, а плод состоит из трех структур, напоминающих фолликулы, за исключением того, что они не совсем отделены друг от друга.
Подсемейство Ochnoideae было пересмотрено в 2014 году как Ochnaceae. Sensu stricto.[3] В этой обработке было описано 27 родов. Дополнительный род, Некия, был воскрешен в том же году на основании результатов молекулярно-филогенетического исследования.[4] Amaral и Bittrich (2014) разделили Ochnoideae на три племени: L Luxembourg- sieae, Sauvagesieae и Ochneae. Род Testulea был включен в племя Sauvagesieae. Подтверждения не распознаны.
В 2014 году была опубликована вторая реклассификация Ochnoideae на основе кладистический анализ Последовательности ДНК. В этой статье Шнайдер и другие разделили Ochnoideae на четыре трибы: Testuleeae, L Luxembourg- siemeae, Sauvagesieae и Ochneae. Их определение племен было таким же, как у Амарала и Биттриха (2014), за исключением того, что Testulea был перемещен из Sauvagesieae в собственное племя Testuleeae. Включение Testulea в Sauvagesieae переводит это племя парафилетический над Люксембургом.
Testulea состоит из одного вида, Testulea gabonensis, что является эндемическим для Габон. Он уникален для Ochnoideae тем, что его листья имеют брохидодромный образец жилкование и его цветы четырехугольный. Кроме того, только одна из тычинок плодородный. Остальные модифицированный в стаминодии и объединенный в столбец до 2⁄3 их длины.
Племя люксембургских состоит из двух родов: Филакра и Люксембург. Филакра является родным для Венесуэла и северный Бразилия. Люксембург из Бразилии.
Пантропическое племя Sauvagesieae состоит из 16 родов, большинство из которых небольшие. Самый большой, безусловно, Sauvagesia, с 38 видами, 35 из которых ограничены неотропы.[11] Sauvagesia неоднороден и может быть парафилетическим даже с Некия снято с него. Отношения в племени Sauvagesieae изучены недостаточно, и по этой причине оно не было разделено на подтяжки.[4]
Племя Ochneae встречается в большинстве тропиков, но наиболее распространено в Африка и тропическая Америка. Он отличается от остальных Ochnoideae абсорбцией эндосперм до того, как семя достигнет зрелость. Его девять родов принадлежат трем подтрибам: Lophirinae, Elvasiinae и Ochninae.[4]
Подтриба Lophirinae состоит из одного рода, Лофира. Имеет два вида, обитающих в тропической Африке. Он дает необычный плод, у которого два чашелистика становятся сильно увеличенный и форма крылья что облегчает распределение семян к ветер.
Подтриба Elvasiinae состоит из двух родов: Периссокарпа и Эльвазия, оба ограничены Американец тропики. Периссокарпа никогда не использовался для молекулярно-филогенетического исследования.
Подтриба Ochninae состоит из шести родов: Campylospermum, Ouratea, Idertia, Brackenridgea, Rhabdophyllum, и Очна. Самый большой из них, Урата, ограничивается Новый мир и содержит все виды Нового Света в Ochninae. Все роды Ochneae кажутся монофилетический, как определено Amaral и Bittrich (2014), но в одном молекулярно-филогенетическом исследовании, Урата и Очна получил только слабый бутстрап поддержка в максимальная вероятность анализ.[4] Idertia и Brackenridgea вероятно, сестринские роды, но никаких других родств среди родов Ochninae не установлено.
Филогения
В филогенетическое дерево ниже адаптировано из опубликованного в 2014 году.[4] Слабо поддерживаемые узлы сворачиваются, образуя политомии. Максимальная вероятность бутстрап поддержка> 75%, если не указано иное. Периссокарпа и Индосиния не были отобраны для ДНК. Их размещение на филогенетическом дереве основано на анатомия и морфология Только.
| MEDUSAGYNOIDEAE QUIINOIDEAE OCHNOIDEAE |
Эволюция
В эволюция of Ochnaceae был необычным, поскольку включал два полных реверсии для актиноморфный цветочная симметрия и два почти полных возврата к апокарпия, состояние, в котором плодолистики полностью отдельные. Актиноморфия и апокарпия считаются "примитивный " состояния персонажей в покрытосеменные.[28] Вторичная апокарпия встречается особенно редко и имеет возникший особенно в Розоцветные, Apocynaceae, Sapindales, и Мальвалес.[12]
Цветки актиноморфны в Медузагайн и Quiinoideae, но у Ochnoideae, зигоморфия это наследственный условие. В Testulea, Филакра, и Люксембург, цветы развивать зигоморфно в бутон. Но в четверке базальный клады of Sauvagesieae, включая роды Blastemanthus, Godoya, Rhytidanthera, Krukoviella, Cespedesia, Fleurydora, Poecilandra, и Wallacea ,, цветы развивать актиноморфно в зародыше, затем становятся зигоморфными после открытие к рост определенных частей цветка. Такая поздняя зигоморфия у цветковых растений встречается очень редко. В оставшейся пятой кладе Sauvagesieae, включающей роды Некия, Schuurmansia, Schuurmansiella, Euthemis, Tyleria, Adenarake, Indosinia, и Sauvagesiaпосле цветения цветки остаются актиноморфными. В трибе Ochneae все виды имеют актиноморфные цветки.
В подтрибе Ochninae и в роде Froesia, компоненты завязи (плодолистики) очень скоро объединенный на базе. В противном случае яичник у Ochnaceae синкарпный, состоящий из полностью оплавленных плодолистиков.
В Медузагайн и Quiinoideae, многие цветки однополые, за исключением Froesia, где они строго гермафродит. У Ochnoideae однополые цветки ограничены кладой, состоящей из Schuurmansia, Schuurmansiella, и Euthemis.[4]
В Медузагайн и Quiinoideae, как и у большинства покрытосеменных, пыльники открываются продольными прорезями. У Ochnoideeae пыльник расхождение является по происхождению порицидный, с несколькими переходами в продольные щели. Testulea, Филакра, и Люксембург есть пыльники, которые открываются апикальный поры. То же самое и с тремя наиболее базальными кладами Sauvagesieae, а именно: Blastemanthus, Флеридора, и кладу из четырех родов с пятью плодолистиками и множеством семяпочек на плодолистик (Годоя, Ритидантера, Круковиелла, и Цеспедезия). Поэчиландра имеет пористое раскрытие пыльника, но в его сестра род, Wallacea, пыльники открываются продольными прорезями.
У остальных Sauvagesieae расхождение пыльников различно. В Schuurmansia, Schuurmansiella, и Аденараке, раскрытие пыльника апикально лонгицидный. Это означает, что продольная щель короткая и не выходит далеко от апикального конца пыльника. У некоторых видов Sauvagesia, пыльники расслаиваются по длине, но весь андроцей завернутый в лепесток стаминодии, таким образом пыльца может вырваться только из верхушки пыльника. Это известно как пористая система, потому что она действует так, как если бы пыльники действительно были пористыми.
У трибы Ochneae расхождение пыльников продольными щелями ограничено Brackenridgea и несколько видов Очна.
Testulea своеобразно иметь только один плодородный тычинка. Его пыльник открывается апикальной порой. Остальные тычинки модифицированный в стерильный стаминодии, объединенные в колонну на срок до 2⁄3 их длины.
В родах Froesia и Quiina, а в племени Ochneae эндосперм полностью всасывается в начале развития семян. Неясно, является ли наличие или отсутствие эндосперма наследственным состоянием Ochnaceae. Долгое время подсемейство Ochnoideae разделялось на две группы только по этому признаку. В такой классификации группа, содержащая эндосперм, будет парафилетический над Очнее, потому что он будет содержать Testulea, Филакра, и Люксембург.
Количество семяпочки на карпель Широко варьирует у Ochnaceae. Медузагайн и Quiinoideae имеют по две семяпочки на плодолистик. В Testulea и в кладе из четырех родов в Sauvagesieae (Годоя, Ритидантера, Круковиелла, и Цеспедезия) количество семяпочек от 100 до 200 на плодолистика. Для остальной части Sauvagesieae, кроме Euthemis, и для Филакра и Люксембургколичество семяпочек на плодолистика колеблется от четырех до 50. Евфемида имеет две семяпочки на плодолистик.
В Очнее, Лофира имеет от 4 до 50 семяпочек на плодолистик. В подтрибах Elvasiinae и Ochninae количество семяпочек на плодолистика равно одной.
История
Семейство Ochnaceae было создано Пирам Августина Кандоль в 1811 г.[29][30] В то время он описал Эльвазия, новый род в семье, и он включил еще трех: Очна, Walkera, и Гомфия.[31] Walkera был описан Иоганн Кристиан Даниэль фон Шребер в 1789 г., но больше не признается. Его типовой вид был описан как Gomphia serrata к Андриас Канис в 1968 г.,[32] но теперь он помещен в род Кампилоспермум.[нужна цитата ] Гомфия давно был источником путаницы[33] и он не был признан в последней редакции Ochnaceae.[3]
Годоя и Sauvagesia были известны в 1811 году, когда де Кандоль основал семью Ochnaceae, но он поместил их в другие семьи. В его Продромус, он поместил Годоя в семье, которая позже будет известна как Clusiaceae.[34] Он считал Лаурадия (Лаврадия) отдельно от Sauvagesia, и поместил их обоих в Violaceae. Он добавил род Castela к Ochnaceae, но теперь он является частью Simaroubaceae.[35] Де Кандоль полагал, что Simaroubaceae был тесно связан с Ochnaceae, но теперь он помещен в порядок Sapindales.[20] Несколько авторов поместили Годоя, Sauvagesia, и другие в семействе Sauvagesiaceae, до начала 21 века.[19] Другие авторы, такие как Адольф Энглер, включил их в Ochnaceae.
В 1874 году Энглер разделил Ochnaceae на две группы на основании отсутствия или наличия эндосперм в зрелом семени.[36] Группа без эндосперма соответствует концепции Ochnaceae де Кандолля и современному трибе Ochneae. Группа с эндоспермом теперь известна как парафилетический и состоит из племен Testuleeae, L Luxembourgieae и Sauvagesieae. В 1876 г. Флора из Бразилия Энглер описал много новых видов Ochnaceae, особенно его самый крупный род, Урата.[37] Он описал 85 видов в Урата, 17 из которых он названный как новый вид в то время. Он также перенес 63 вида в Урата от других родов.
Роды Quiina и Touroulia были известны с 1775 г., когда они были описаны Жан Батист Обле,[38] и они были по-разному классифицированы по 19 веку систематики. Жак Дени Шуази построил для них семейство Quiinaceae (как Quiinacées) в 1849 году,[39] но он не встретил требования за действующая публикация из ботаническое название. Название Quiinaceae было подтверждено Энглером в Флора Бразилиенсис в 1888 г.[30][40]
Род Medusagyne был описан Джон Гилберт Бейкер в 1877 г. во флоре Маврикий и Сейшельские острова,[41] но только в 1924 году он был выделен в отдельную моногенную семью.[42]
В 1893 г. Эрнест Фридрих Гильг покрытые Ochnaceae и Адольф Энглер покрыл Quiinaceae для первого издания Die Natürlichen Pflanzenfamilien.[43][44] Энглер написал описание Медузагайн в добавка к первому версия из DNP в 1897 г.[45] Он разместил Медузагайн под заголовком «Zweifelhafte, möglicherweise zu den Guttiferae gehörige Gattung» (сомнительный, возможно, род, принадлежащий к Guttiferae). Guttiferae - это устаревший имя для Clusiaceae.
В 1902 г. Филипп ван Тигем выделили шесть семейств в подсемействе Ochnoideae.[46] Это были люксембургские, соважные, обыкновенные, эвтемидовые, лофирские и очковые. Три из них (Wallaceae, Euthemidaceae и Lophiraceae) были моногенными и были возведены ван Тигхемом в то время. Его люксембургские включали базальную клады из того, что сейчас является племенем Sauvagesieae. Ван Тигем назвал очень много родов в 1902 году, ограничивая их очень узко. В том, что сейчас подкуп Ochninae выделил 53 рода. Самая последняя редакция этой группы делит ее на шесть родов.
В 1925 г. для второго издания DNP, Энглер и Гилг расширили свои лечение Quiinaceae и Ochnaceae, соответственно, по сравнению с тем, что они написали в 1893 году.[47][48] Medusagynaceae была покрыта в том же объеме DNP Адольф Энглер и Ганс Мельхиор.[49]
Для второго издания DNP, Энглер распознал два рода, Quiina и Touroulia, у Quiinaceae. Лакунария и Froesia были обнаружены позже и названы в 1925 и 1948 годах соответственно.
В том же томе Гилг разделил Ochnaceae (эквивалент современных Ochnoideae) на 21 род, включая Индоветия, Лейтгебия, Ваусагесия, и Лаурадия (в качестве Лаврадия), которые теперь рассматриваются как синонимы из Sauvagesia.[3] Восемь современных родов, (Филакра, Круковиелла, Флеридора, Тилерия, Аденараке, Индосиния, Периссокарпа, и Idertia) состоят из растений, которые не были обнаружены ботаническое исследование в это время. Гилг разместил Ритидантера в синонимии под Годоя, но Ритидантера принято сегодня. Он включил Кампилоспермум и Рабдофиллум в Гомфия и разместил Гомфия в синонимии под Урата. В своей редакции Ochnaceae Гилг представил резюме классификации ван Тигема, а также его собственная.[48] Три из родов ван Тигема, (Кампилоспермум, Рабдофиллум, и Ритидантера) все еще признаны сегодня.[3]
В 1968 г. Андриас Канис опубликовал бумага это сильно повлияло на последующую работу по Ochnaceae, вплоть до редакции 2014 года.[32] Клод Х.Л. Састре назвал много новых видов Ochnaceae, в нескольких документы с 1970 по 2003 гг.[4]
В 1991 г. кладистический был опубликован анализ Ochnaceae.[50] В том же году Некия, 28-й род Ochnoideae, был возрожден в молекулярный филогенетический исследование на основе четырех хлоропластная ДНК места, и ядерный рибосомальный ЭТО.[4] Семьдесят девять видов Ochnaceae были отобранный и была представлена новая классификация. Также, Testulea был исключен из племени Sauvagesieae и помещен в монотипный племя Testuleeae.
Рекомендации
- ^ а б c Группа филогении покрытосеменных (2009). «Обновленная классификация филогенетической группы покрытосеменных для порядков и семейств цветковых растений: APG III» (PDF). Ботанический журнал Линнеевского общества. 161 (2): 105–121. Дои:10.1111 / j.1095-8339.2009.00996.x. Получено 2013-07-06.
- ^ а б c Вернон Х. Хейвуд, Ричард К. Браммит, Оле Себерг и Аластер Калхэм. Семейства цветущих растений мира. Firefly Books: Онтарио, Канада. (2007). ISBN 978-1-55407-206-4.
- ^ а б c d е ж грамм час я j k Мария ду Карму Э. Амарал и Фолькер Биттрих. 2014. «Ochnaceae». страницы 253-268. Дои:10.1007/978-3-642-39417-1_19 В: Клаус Кубицки (редактор). 2014 г. Семейства и роды сосудистых растений том XI. Springer-Verlag: Берлин, Гейдельберг, Германия. ISBN 978-3-642-39416-4 (Распечатать). ISBN 978-3-642-39417-1 (электронная книга). Дои:10.1007/978-3-642-39417-1
- ^ а б c d е ж грамм час я j k л м п о п q Хулио В. Шнайдер, Пульчери Биссиенгу, Мария ду Карму Э. Амарал, Али Тахир, Майкл Ф. Фэй, Марко Тинес, Марк С. Сосеф, Георг Жижка и Ларс В. Чатру. 2014. «Филогенетика, реконструкция предкового состояния и новая инфрасемейная классификация пантропических Ochnaceae (Medusagynaceae, Ochnaceae s.str., Quiinaceae) на основе пяти областей ДНК». Молекулярная филогенетика и эволюция 78:199-214. Дои:10.1016 / j.ympev.2014.05.018.
- ^ а б c Чжэнсян Си, Брэд Р. Рухфель, Ханно Шефер, Андре М. Аморим, Маникам Сугумаран, Кеннет Дж. Вурдак, Питер К. Эндресс, Мерран Л. Мэтьюз, Питер Ф. Стивенс, Сара Мэтьюз и Чарльз К. Дэвис. 2012. "Филогеномика и апостериорное разделение данных решают меловые покрытосеменные радиации Malpighiales". PNAS (Труды Национальной академии наук Соединенных Штатов Америки) 109(43):17519-17524. Дои:10.1073 / pnas.1205818109. (Видеть внешняя ссылка ниже).
- ^ а б Кеннет Дж. Вурдак; Чарльз С. Дэвис (2009), «Филогенетика Malpighiales: завоевание позиций на одной из самых непокорных клад на дереве жизни покрытосеменных», Американский журнал ботаники, 96 (8): 1551–1570, Дои:10.3732 / ajb.0800207, PMID 21628300
- ^ Christenhusz, M. J. M .; Бинг, Дж. У. (2016). «Количество известных видов растений в мире и его ежегодный прирост». Фитотакса. 261 (3): 201–217. Дои:10.11646 / phytotaxa.261.3.1.
- ^ Поль А. Де Лука и Марио Вальехо-Марин. 2013. «О чем идет шум? Экология и эволюционное значение опыления кайфом». Текущее мнение в области биологии растений 16(4):429-435. Дои:10.1016 / j.pbi.2013.05.002.
- ^ Энтони Хаксли, Марк Гриффитс и Марго Леви (1992). Словарь садоводства нового Королевского садоводческого общества. Macmillan Press, Limited: Лондон. Стоктон Пресс: Нью-Йорк. ISBN 978-0-333-47494-5 (набор).
- ^ Уоррен Л. Вагнер, Деррал Р. Хербст и Сай Х. Зомер. Руководство цветущих растений Гавайев, Исправленное издание, 1999. Издательство Епископского музея: Hololulu
- ^ а б Дэвид Дж. Мабберли. 2008 г. Книга растений Мабберли третье издание (2008 г.). Издательство Кембриджского университета: Великобритания. ISBN 978-0-521-82071-4 (Видеть внешняя ссылка ниже).
- ^ а б Питер К. Эндресс. 2011. «Эволюционное разнообразие цветков у покрытосеменных растений». Американский журнал ботаники 98(3):370-396. Дои:10.3732 / ajb.1000299. (Видеть внешняя ссылка ниже).
- ^ Дэниел Данехи, Питер Уилф и Стефан А. Литтл. 2007. «Макрофлора раннего эоцена из места остановки красных горячих грузовиков (Меридиан, Миссисипи, США)». Palaeontologia Electronica 10(3): 17A: 31 стр. (Видеть внешняя ссылка ниже).
- ^ Сусана Магаллон, Хидир В. Хилу и Дитмар Квандт. 2013. «Хронология эволюции наземных растений: эффекты генов вторичны по сравнению с ограничениями на ископаемые в нестрогой временной оценке возраста и темпов замещения». Американский журнал ботаники 100(3):556-573. Дои:10.3732 / ajb.1200416. (Видеть внешняя ссылка ниже).
- ^ Список растений: Ochnaceae. (Видеть внешняя ссылка ниже).
- ^ а б Хулио В. Шнайдер, Ульф Свенсон, Розабель Самуэль, Тод Стюесси и Георг Жижка. 2006. «Филогенетика Quiinaceae (Malpighiales): данные о последовательности и морфологии trnL-trnF». Систематика и эволюция растений 257(3-4):189-203. Дои:10.1007 / s00606-005-0386-5.
- ^ а б Клаус Кубицки. 2014. «Quiinaceae». страницы 277-281. Дои:10.1007/978-3-642-39417-1_22. В: Клаус Кубицки (редактор). 2014 г. Семейства и роды сосудистых растений том XI. Springer-Verlag: Берлин, Гейдельберг, Германия. ISBN 978-3-642-39416-4 (Распечатать). ISBN 978-3-642-39417-1 (электронная книга). Дои:10.1007/978-3-642-39417-1
- ^ а б Уильям К. Дикисон и Клаус Кубицки. 2014. «Medusagynaceae». страницы 249-251. Дои:10.1007/978-3-642-39417-1_18. В: Клаус Кубицки (редактор). 2014 г. Семейства и роды сосудистых растений том XI. Springer-Verlag: Берлин, Гейдельберг, Германия. ISBN 978-3-642-39416-4 (Распечатать). ISBN 978-3-642-39417-1 (электронная книга). Дои:10.1007/978-3-642-39417-1
- ^ а б Армен Львович Тахтаджан (Тахтаджян). Цветущие растения второе издание (2009 г.). Springer Science + Business Media. ISBN 978-1-4020-9608-2. ISBN 978-1-4020-9609-9 Дои:10.1007/978-1-4020-9609-9. (Видеть внешняя ссылка ниже).
- ^ а б c d Питер Ф. Стивенс (с 2001 г.). "Ochnaceae" по адресу: Сайт филогении покрытосеменных. В: Ботанический сад Миссури Интернет сайт. (видеть внешняя ссылка ниже)
- ^ Клеменс Байер. 2003 г. Дигодендрон. страницы 175-177. В: Клаус Кубицки (редактор). 2003 г. Семейства и роды сосудистых растений Том V. (редакторы тома: К. Кубицки и К. Байер). Springer-Verlag: Берлин, Гейдельберг, Германия. ISBN 978-3-540-42873-2 (Распечатать) ISBN 978-3-662-07255-4 (электронная книга). Дои:10.1007/978-3-662-07255-4.
- ^ Питер Ф. Стивенс. 2007. «Clusiaceae - Guttiferae». страницы 48-66. В: Клаус Кубицки (редактор). 2007 г. Семейства и роды сосудистых растений том IX. (редакторы тома: К. Кубицки в сотрудничестве с К. Байером и П.Ф. Стивенсом). Springer-Verlag: Берлин, Гейдельберг, Германия. ISBN 978-3-540-32214-6 () ISBN 978-3-540-32219-1 () Дои:10.1007/978-3-540-32219-1.
- ^ Брэд Р. Рухфель, Питер Ф. Стивенс и Чарльз С. Дэвис. 2013. «Комбинированная морфологическая и молекулярная филогения кластероидной клады (Malpighiales) и размещение древних розидных макрофоссилий. Палеоклюзия". Международный журнал наук о растениях 174(6):910–936. Дои:10.1086/670668.
- ^ Питер К. Эндресс, Чарльз С. Дэвис и Мерран Л. Мэтьюз. 2013. «Достижения в характеристике цветочной структуры основных субкладов Malpighiales, одного из крупнейших отрядов цветковых растений». Анналы ботаники 111(5): 969–985. Дои:10.1093 / aob / mct056. (Видеть внешняя ссылка ниже).
- ^ Мерран Л. Мэтьюз, Мария ду Карму Э. Амарал и Питер К. Эндресс. 2012. «Сравнительная цветочная структура и систематика у Ochnaceae s.l. (Ochnaceae, Quiinaceae и Medusagynaceae; Malpighiales)». Ботанический журнал Линнеевского общества 170(3):299-392. Дои:10.1111 / j.1095-8339.2012.01299.x.
- ^ Уильям С. Дикисон. 1990. «Морфология и взаимоотношения Медузагайн (Medusagynaceae) ". Систематика и эволюция растений 171(1-4):27-55.
- ^ Хулио В. Шнайдер и Георг Жижка. в прессе. «Quiinaceae». Флора Neotropica монография ??.
- ^ Дуглас Э. Солтис, Памела С. Солтис, Питер К. Эндресс и Марк У. Чейз. 2005 г. Филогения и эволюция покрытосеменных растений. Sinauer Associates, Inc.: Сандерленд, Массачусетс, США. ISBN 978-0-87893-817-9.
- ^ Ochnaceae в Международный указатель названий растений. (видеть внешняя ссылка ниже).
- ^ а б Джеймс Л. Ривил. 2008 г. «Контрольный список семейных и надсемейных названий для существующих сосудистых растений». В: Домашняя страница Джеймса Л. Ривила и К. Роуз Брум. (видеть внешняя ссылка ниже).
- ^ Пирам Августина Кандоль. 1811 г. Nouveau Bulletin des Sciences par la Société Philomathique de Paris 2 (40): 208. (Видеть внешняя ссылка ниже).
- ^ а б Андриас Канис. 1968. «Пересмотр Ochnaceae Индо-Тихоокеанского региона». Blumea 16(1):1-83.
- ^ Вернер Гройтер и Роза Ранкин Родригес. 2014. «Типа для Гомфия (Ochnaceae) - еще раз ". Таксон 63(5):1122-1123. Дои:10.12705/635.5
- ^ Пирам Августина Кандоль. 1824. «Violaceae» (как Violarieae), страницы 287-316; Clusiaceae (как Guttiferae), страницы 557-564; «Ochnaceae», страницы 735-738. В: Prodromus Systematis Naturalis Regni Vegetabilis том 1. (см. внешняя ссылка ниже).
- ^ Джошуа В. Клейтон. 2011. "Simaroubaceae", страницы 408-423. В: Клаус Кубицки (редактор). 2011 г. Семейства и роды сосудистых растений том X. Springer-Verlag: Берлин, Гейдельберг, Германия. ISBN 978-3-642-14396-0 (Распечатать). ISBN 978-3-642-14397-7 (электронная книга). Дои:10.1007/978-3-642-14397-7
- ^ Его Величество Адольф Энглер. 1874. "Ueber Begrenzung und systematische Stellung der natürlichen Familie der Ochnaceae". Nova Acta Academieae Caesarieae Leopoldino - Carolinae Germanicae Naturae Curiosorum 37(2): 1-28. (Видеть внешняя ссылка ниже).
- ^ Его Величество Адольф Энглер. 1876. «Ochnaceae» В: Карл Ф.П. фон Мартиус и Август Вильгельм Эйхлер (редакторы). Флора Бразилиенсис 12(2): 297-366. (Видеть внешняя ссылка ниже).
- ^ Жан Батист К.Ф. Aublet. 1775. Histoire des Plantes de la Guiane Françoise: · · · . Quiina: Приложение к томам 1 и 2, стр. 19; Touroulia: 1:492; Урата: 1: 397. (Видеть внешняя ссылка ниже).
- ^ Жак Дени Шуази. 1849 г. Описание Guttiferes de l'Indie, · · · :12.
- ^ Его Величество Адольф Энглер. 1888. «Quiinaceae». страницы 475-486 и таблицы 109-110. В: Карл Ф.П. фон Мартиус и Август Вильгельм Эйхлер (редакторы). Флора Бразилиенсис 12(1). (Видеть внешняя ссылка ниже).
- ^ Джон Гилберт Бейкер. 1877 г. Флора Маврикия и Сейшельских островов: 16. Л. Рив и Ко .: Лондон, Великобритания. (Видеть внешняя ссылка ниже).
- ^ Г. Г. Адольф Энглер и Эрнест Ф. Гилг. 1924 г. Syllabus der Pflanzenfamilien: eine Übersicht · · · neunte und zehnte auflage: 280. (Программа по семействам растений: обзор · · · выпуски 9 и 10, стр. 280.)
- ^ Эрнест Ф. Гилг. 1893. «Ochnaceae». страницы 131-153. В: Герберт Адольф Энглер и Карл А.Е. Прантл (редакторы). 1895 г. Die Natürlichen Pflanzenfamilien 1-е издание: Teil III, Abteilung 6 (том 3, часть 6). Verlag von Wilhelm Engelmann: Лейпциг, Германия. (Видеть внешняя ссылка ниже).
- ^ Его Величество Адольф Энглер. 1893. «Quiinaceae». страницы 165-167. В: Герберт Адольф Энглер и Карл А.Е. Прантл (редакторы). 1895 г. Die Natürlichen Pflanzenfamilien 1-е издание: Teil III, Abteilung 6 (том 3, часть 6). Verlag von Wilhelm Engelmann: Лейпциг, Германия. (Видеть внешняя ссылка ниже).
- ^ Его Величество Адольф Энглер. 1897 г. Медузагайн. стр. 250. В: Герберт Адольф Энглер. 1897. «Guttiferae». страницы 247-250. В: HG Адольф Энглер и Карл А.Э. Прантл. 1897 г. Die Natürlichen Pflanzenfamilien: Nachträge [I] zum II-IV Teil. (Природные семейства растений: первое приложение к томам 2–4). (Видеть внешняя ссылка ниже).
- ^ Филипп Э. ван Тигем. 1902. «Sur les Ochnacées». Анналы естественных наук - ботаника, серия 8 16:161-416.
- ^ Его Величество Адольф Энглер. 1925. «Quiinaceae». страницы 106-108. В: Герберт Адольф Энглер и Карл А.Е. Прантл (редакторы). 1925 г. Die Natürlichen Pflanzenfamilien: zweite auflage 21 Band. (Природные семейства растений: 2-е издание, том 21). Verlag von Wilhelm Engelmann: Лейпциг, Германия.
- ^ а б Эрнест Ф. Гилг. 1925. «Ochnaceae». страницы 53-87. В: Герберт Адольф Энглер и Карл А.Е. Прантл (редакторы). 1925 г. Die Natürlichen Pflanzenfamilien: zweite auflage 21 Band. (Природные семейства растений: 2-е издание, том 21). Verlag von Wilhelm Engelmann: Лейпциг, Германия.
- ^ Ганс Мельхиор. 1925. «Medusagynaceae». страницы 50-52. В: Герберт Адольф Энглер и Карл А.Е. Прантл (редакторы). 1925 г. Die Natürlichen Pflanzenfamilien: zweite auflage 21 Band. (Природные семейства растений: 2-е издание, том 21). Verlag von Wilhelm Engelmann: Лейпциг, Германия.
- ^ Мария ду Карму Э. Амарал. 1991. "Phylogenetische Systematik der Ochnaceae". Botanische Jahrbücher für Systematik, Pflanzengeschichte, und Pflanzengeographie 113(1):105-196.
'
внешняя ссылка
- Xi2012 В: Pubmed Central В: NCBI (Национальный центр биотехнологической информации)
- Цеспедезия И Очна И Sauvagesia В Книга растений Мабберли
- Эндресс (2011) И хронология эволюции наземных растений В: Архив В: Американский журнал ботаники
- Эоценовая макрофлора из Миссисипи (печать) В: оглавление В: том 10, выпуск 3 В: задние выпуски по обложке В: Обзор статей В: Palaeontologia Electronica/содержание
- Ochnaceae В: покрытосеменные В: Просматривать В: Список растений
- Ochnaceae В Тахтаджан 2009 г.
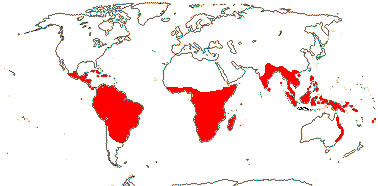
- Карта распространения Ochnoideae ИСписок родов Ochnoideae В Ochnoideae ВМальпигиалес В Деревья В: APweb В: Ботанический сад Миссури
- кластероидная клада и Палеоклюзия В: том 174, выпуск 6 Из: IJPS
- цветочная структура в Мальпигиалес В: Архив В: Анналы ботаники
- Ochnaceae В: Названия растений В: IPNI
- OA-OZ И Q В Контрольный список надродовых названий сохранившихся сосудистых растений В Домашняя страница Джеймса Л. Ривила и К. Роуз Брум
- Гомфия В Таксон
- Флора Маврикия и Сейшельских островов В Archive.org
- В BHL (Библиотека наследия биоразнообразия ):
{kind=link}
- Ochnaceae, стр.208 В Выпуск 40 В Просмотр книги В Nouveau Bulletin des Sciences par la Société philomathique de Paris, том 2
- Лаврадия И Sauvagesia И Годоя И Castela В Просмотр книги В Продромус т.1 (1824 г.)
- Ochnaceae том 37, номер 2 В том 37 В Nova acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum
- Ochnaceae В volume12 часть 2 В Флора Бразилиенсис
- Quiina В Дополнение к тому 2 В том 2 ИTouroulia И Урата В том 1 Из Histoire des planttes de la Guiane Françoise
- Quiinaceae В volume12 часть 1 В Флора Бразилиенсис
- Ochnaceae И Quiinaceae В Том 3 часть 6 Из Die Natürlichen Pflanzenfamilien
- Медузагайн В Nachträge (Дополнение) Teil 2-4 В Die Natürlichen Pflanzenfamilien
- Sur les Ochnacées В Серия 8, том 16 Из Annales des Sciences naturelles, huitieme série - Botanique.
- Ochnaceae (точный поиск) В: Имена В: Тропикос В: Наука и сохранение В: Ботанический сад Миссури
- Ochnaceae в Л. Уотсон и М.Дж. Даллвиц (с 1992 г.). Семейства цветковых растений: описания, иллюстрации, идентификация, поиск информации. http://delta-intkey.com
- Список родов Ochnaceae В: Двудольные В: Список родов в семье В: Семейства и роды сосудистых растений В: О контрольном списке В: Всемирный контрольный список избранных семейств растений В: Названия растений В: Источники данных В: ePIC В: Базы данных В: Ресурсы и базы данных В: Научные исследования и данные В: Наука и охрана В: Kew Gardens
- Неотропические Ochnaceae В: Семейный индекс В: Неотропики В: Проекты и программы В: Проект Тропическая Америка В: Кью в глубине В: Научные исследования и данные В: Kew Gardens
- Ochnaceae В: Мальпигиалес В: Магнолиопсида В: Трахеофита В: Растения В: Глобальные виды
- Ochnaceae В: Покрытосеменные В: Просматривать В: Список растений
- Показать результаты В: Иерархический отчет с выбранными Plantae и Genus В: Доступ к данным В: ЭТО
- Мальпигиалес В: розиды В: основные эвдикоты (Gunneridae) В: эвдикоты В: покрытосеменные В: Сперматопсиды В: эмбриофиты В: зеленые растения В: эукариоты В: жизнь на земле (корень дерева) В: Веб-проект "Древо жизни" (ToL)
- Полный список родов В: Ochnaceae В: Список семей В: Семьи и роды в GRIN В: Запросы В: Таксономия GRIN для растений
- Ochnaceae В: Мальпигиалес В: сказочные В: розиды В: Пентапеталы В: Gunneridae В: евдикотиледоны В: Мезангиоспермия В: Magnoliophyta (цветковые растения) В: Сперматофиты В: Euphyllophyta В: Трахеофита В: Эмбриофита В: Стрептофитина В: Streptophyta В: Viridiplantae В: Эукариоты В: Таксономия В: UniProt
- Ochnaceae В: Мальпигиалес В: сказочные В: розиды В: Пентапеталы В: Gunneridae В: евдикотиледоны В: Мезангиоспермия В: Magnoliophyta (цветковые растения ··· В: ··· Эукариоты В: Браузер таксономии В: База данных таксономии В: Таксономия В: NCBI (Национальный центр биотехнологической информации)