Equisetidae - Equisetidae
| Equisetidae | |
|---|---|
 | |
| Equisetum telmateia | |
| Научная классификация | |
| Королевство: | Plantae |
| Clade: | Трахеофиты |
| Учебный класс: | Полиподиопсиды |
| Подкласс: | Equisetidae Теплый. |
| Заказы | |
| Синонимы | |
См. Текст. | |
Equisetidae один из четырех подклассы из Полиподиопсиды (папоротники), группа сосудистые растения с летописью окаменелостей, восходящей к Девонский. Они широко известны как хвощи.[2] Обычно они растут на влажных участках, с мутовками игольчатых ветвей, расходящихся через равные промежутки времени от одного вертикального стебля.
Раньше Equisetidae считались отдельным разделение споровых растений и называется Equisetophyta, Артрофит, Calamophyta или же Sphenophyta. Когда рассматриваются как класс, имена Equisetopsida SS. и Sphenopsida также использовались. Теперь они признаны довольно близкими родственниками папоротники (Polypodiopsida), из которых они образуют специализированную ветвь.[3] Однако разделение на хвощи и другие папоротники настолько древнее, что многие ботаники, особенно палеоботаники, по-прежнему считают эту группу принципиально отдельной на более высоком уровне.
Описание
Хвощи содержат фотосинтезирующие, «сегментированные» полые стебли, иногда наполненные сердцевиной. На стыке («узел», см. Диаграмму) между каждым сегментом находится завиток листья. В единственном дошедшем до нас роде Equisetum, это маленькие листочки (микрофиллы ) с единичным сосудистым следом, сросшимся в оболочку на каждом стеблевом узле. Однако листья Equisetum вероятно возникла в результате сокращения мегафиллы, о чем свидетельствуют ранние ископаемые формы, такие как Sphenophyllum, у которого листья широкие с ветвистыми жилками.[4]
В сосудистые пучки тройчатые в узлах, при этом центральная ветвь становится жилкой микрофилла, а две другие перемещаются влево и вправо, чтобы слиться с новыми ветвями своих соседей.[5] Сама сосудистая система напоминает сосудистую систему растений. Eustele, которые развивались независимо и конвергентно.[5] Очень быстрое удлинение междоузлий приводит к образованию пробковой полости и кольца каринальные каналы образуется в результате нарушения первичного ксилема. Подобные пространства, валлекулярные каналы образуются в коре головного мозга.[5] Из-за более мягкой природы флоэмы они очень редко встречаются в ископаемых экземплярах.[нужна цитата ] в Calamitaceae, вторичная ксилема (но не вторичная флоэма ) был секретен как камбий росла наружу, давая древесный стебель и позволяя растениям вырастать до 10 метров. Все существующие виды Equisetum травянистые и утратили способность давать вторичный рост.[5]
Подземные части растений состоят из сочлененных корневища, из которых выходят корни и надземные оси. У растений есть вставочные меристемы в каждом сегменте стебля и корневища, которые растут по мере роста растения. Это контрастирует с семенными растениями, которые растут из апикальной меристемы - то есть новый рост происходит только от кончиков роста (и расширения стеблей).
Хвощи медвежьи шишки (технически стробили, петь. стробил ) на кончиках некоторых стеблей. Эти конусы содержат спирально расположенные спорангиеносцы, которые несут спорангии по краям, а у сохранившихся хвощей покрывают споры внешне - как мешочки, свисающие с зонта, ручка которого расположена на оси конуса. В вымерших группах дополнительную защиту спорам обеспечивало присутствие мутовок. прицветники - большие заостренные микрофиллы, выступающие из конуса.
Сохранившиеся хвощи гомоспористый, но вымершие гетероспористый такие виды, как Calamostachys casheana появляются в летописи окаменелостей.[6] Спорангии открываются боковыми расхождение чтобы выпустить споры. Споры несут характерный Elaters, отличительные пружинные насадки, которые гигроскопичный: то есть они меняют свою конфигурацию в присутствии воды, помогая спорам двигаться и способствуя их распространению.
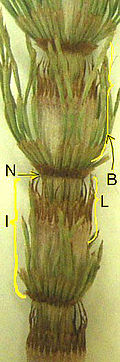
N = узел,
I = междоузлия,
B = ветвь в мутовке,
L = слитые микрофиллы


Таксономия
Классификация
Хвощи и их ископаемые родственники долгое время считались отличными от других бессемянных. сосудистые растения, такой как папоротники (Полиподиопсиды).[7] До появления современных молекулярные исследования, связь этой группы с другими живыми и ископаемыми растениями считалась проблематичной.[8] Из-за их неясных отношений звание, присвоенное ботаниками хвощам, варьировалось от порядок к разделение. При выделении в отдельный раздел в литературе используется множество возможных названий, включая Arthrophyta,[8] Calamophyta, Sphenophyta,[1][9] или Equisetophyta. Другие авторы рассматривали ту же группу как класс либо внутри подразделения, состоящего из сосудистых растений, либо, в последнее время, внутри расширенной группы папоротников. При расчете на класс группа получила название Equisetopsida.[10] или Sphenopsida.[5]
Современное филогенетический анализ еще в 2001 году продемонстрировали, что хвощ прочно входит в папоротник клады из сосудистые растения.[11][12] Smith et al. (2006) провели первую высокоуровневую птеридофит классификация опубликована в молекулярный филогенетический эры, и считалось, что папоротники (монилофиты) составляли четыре классы, с хвощами как класс Equisetopsida Sensu stricto.[3] (Это различие необходимо из-за альтернативного использования Equisetopsida Sensu lato как синоним всех наземных растений (Embryophyta) с рангом класса.[13]) гнаться и Раскрыть (2009) рассматривали хвощи как подкласс Equisetidae класса Equisetopsida. Sensu lato. Консенсусная классификация, разработанная Филогенетической группой Pteridophyte в 2016 году, также помещает хвощи в подкласс Equisetidae, но в класс Polypodiopsida (папоротники в широком смысле).[2]
Филогения
На следующей диаграмме показана вероятная филогенная взаимосвязь между подклассом Equisetidae и другими подклассами папоротников в соответствии с Филогенетической группой Pteridophyte.[2]
| Полиподиопсиды |
| ||||||||||||||||||
Исследование 2018 г., проведенное Elgorriaga et al. предполагает, что отношения внутри Equisetidae показаны на следующей кладограмме.[14]
| |||||||||||||||||||||||||||||||
Согласно исследованию, возраст коронной группы Equisetum дат по крайней мере до Раннемеловой период, и, скорее всего, до Юрский.[14]
Подразделение
Подкласс Equisetidae содержит единственный существующий отряд, Equisetales. Этот заказ состоит из одного монотипный семья, Хвойные, с одним родом Equisetum. Equisetum насчитывает около 20 видов.[12][2]
Окаменелости
Современные хвощи представляют собой лишь небольшую часть разнообразия хвощей прошлого. Было три отряда Equisetidae. В Pseudoborniales впервые появился в конце Девонский.[1] В Sphenophyllales были доминирующим членом Каменноугольный подлеска, и процветал до середины и ранняя пермь. В Equisetales существовал рядом с Sphenophyllales, но разнообразили, поскольку эта группа исчезла, постепенно уменьшаясь в разнообразии до сегодняшнего единственного рода Equisetum.
Эти организмы впервые появляются в летописи окаменелостей в конце девона,[1] время, когда наземные растения подвергались быстрой диверсификации, когда корни, семена и листья только развивались. (Видеть Эволюционная история растений ) Однако растения существовали на суше уже почти сто миллионов лет, и первые свидетельства того, что наземные растения датируются 475 миллион лет назад.[15]
Рекомендации
- ^ а б c d Тейлор, Т. И Taylor., E.L. (1993). Биология и эволюция ископаемых растений. Энглвуд Клиффс, Нью-Джерси: Prentice Hall. С. 303–305. ISBN 978-0-13-651589-0.
- ^ а б c d Pteridophyte Phylogeny Group I (ноябрь 2016 г.), «Классификация современных ликофитов и папоротников на основе сообществ», Журнал систематики и эволюции, 54 (6): 563–603, Дои:10.1111 / jse.12229, S2CID 39980610CS1 maint: ref = harv (связь)
- ^ а б Smith, A.R .; Прайер, К.М .; Schuettpelz, E .; Korall, P .; Шнайдер, Х. и Вольф, П.Г. (2006). «Классификация сохранившихся папоротников» (PDF). Таксон. 55 (3): 705–731. Дои:10.2307/25065646. JSTOR 25065646. Архивировано из оригинал (PDF) на 2008-02-26. Получено 2019-10-28.
- ^ Rutishauser, R. (1999). «Полимерные мутовки листьев сосудистых растений: морфология развития и нечеткость органов». Международный журнал наук о растениях. 160 (6): 81–103. Дои:10.1086/314221. JSTOR 10.1086/314221. PMID 10572024.
- ^ а б c d е Стюарт, W.N. & Rothwell, G.W. (1993). Палеоботаника и эволюция растений (2-е изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-38294-6.
- ^ Бауэр, Ф. (1959) [1935]. Происхождение наземной флоры, теория, основанная на фактах чередования. Нью-Йорк: Hafner Publishing Co., стр. 381.
- ^ Имс, А.Дж. (1936). Морфология сосудистых растений (нижние группы). Нью-Йорк и Лондон: Книжная компания Макгроу-Хилл. С. 110–115.
- ^ а б Bold, H.C .; Alexopoulos, C.J .; Делеворяс, Т. (1987). Морфология растений и грибов (5-е изд.). Нью-Йорк: Харпер-Коллинз. С. 371–387, 478, 506–514. ISBN 978-0-06-040839-8.
- ^ Gifford, E.M .; Фостер, А. (1988). Морфология и эволюция сосудистых растений (3-е изд.). Нью-Йорк: В. Х. Фриман и компания. стр.175 –207. ISBN 978-0-7167-1946-5.
- ^ Kenrick, P .; Крейн, П.Р. (1997). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование. Вашингтон, округ Колумбия: Пресса Смитсоновского института. С. 241–242. ISBN 978-1-56098-730-7.
- ^ Прайер, К.М .; Schneider, H .; Smith, A.R .; Cran, R .; Wolf, P.G .; Хант, J.S .; Сайпс, С. (2001). «Хвощи и папоротники - монофилетическая группа и ближайшие живущие родственники семенных растений» (PDF). Природа. 409 (6820): 618–621. Bibcode:2001Натура.409..618S. Дои:10.1038/35054555. PMID 11214320. S2CID 4367248. Архивировано из оригинал (PDF) на 2009-09-02.
- ^ а б Кристенхуц, Маартен Дж. М. и Чейз, Марк В. (2014). «Тенденции и концепции классификации папоротников». Анналы ботаники. 113 (4): 571–594. Дои:10.1093 / aob / mct299. ЧВК 3936591. PMID 24532607.CS1 maint: ref = harv (связь)
- ^ Чейз, Марк В. & Открой, Джеймс Л. (2009), «Филогенетическая классификация наземных растений, сопровождающих APG III», Ботанический журнал Линнеевского общества, 161 (2): 122–127, Дои:10.1111 / j.1095-8339.2009.01002.xCS1 maint: ref = harv (связь)
- ^ а б Эльгорриага, А .; Escapa, I.H .; Rothwell, G.W .; Tomescu, A.M.F .; Кунео, Н. (2018). "Происхождение Equisetum: эволюция хвоща (Equisetales) в пределах основной клады эвфиллофитов Sphenopsida". Американский журнал ботаники. 105 (8): 1286–1303. Дои:10.1002 / ajb2.1125. PMID 30025163.
- ^ Wellman, C.H .; Osterloff, P.L .; Мохиуддин, У. (2003). «Фрагменты древнейших наземных растений» (PDF). Природа. 425 (6955): 282–285. Bibcode:2003 Натур. 425..282Вт. Дои:10.1038 / природа01884. PMID 13679913. S2CID 4383813.
![]() Данные, относящиеся к Equisetopsida в Wikispecies
Данные, относящиеся к Equisetopsida в Wikispecies
| Викискладе есть медиафайлы по теме Equisetopsida. |